Pubblicato il 08.03.19
L’apparato radicale del riso si compone diverse tipologie di radici che differiscono tra loro per struttura istologica e anatomica, nonché per funzione. Si distinguono radici dette seminali che originano dall’embrione del seme e che degenerano rapidamente, della corona che si formano progressivamente a partire da ciascuno dei nodi alla base del culmo e, infine, avventizie che si differenziano a partire dai nodi aerei. Indipendentemente dall’origine da ciascuna radice possono svilupparsi radici secondarie. Il numero, la lunghezza e l’angolo di crescita di questo insieme di radici costituiscono l’architettura dell’apparato radicale, un tratto morfo-funzionale stabilito da determinati genetiche e dalla loro interazione con i fattori ambientali, inclusi quelli che si instaurano nel suolo come conseguenza di agrotecniche specifiche.
Architetture radicali che privilegiano lo sviluppo delle singole radici negli strati più superficiali del suolo favoriscono l’acquisizione dei nutrienti minerali meno mobili, mentre architetture più profonde garantiscono un’efficiente acquisizione di acqua e dei nutrienti minerali più mobili (Fig. 1). Quindi le cultivar di riso caratterizzate da architetture radicali più profonde sono più adatte di altre a condizioni di bassa disponibilità di azoto nitrico, mentre quelle con architetture radicali più superficiali sono adatte a suoli con scarsa disponibilità di fosfati.
Fra le variabili ambientali che più di altre influenzano l’architettura radicale delle piante vanno considerate le proprietà fisiche (tessitura, compattezza, densità, porosità, permeabilità), chimiche (pH, pressione parziale di ossigeno, dotazione di sostanza organica, disponibilità di elementi minerali nutritivi) e microbiologiche (microrganismi rizosferici promotori di crescita, simbionti o patogeni) del suolo. Ne consegue che l’efficienza complessiva in risicoltura di agrotecniche, come il sovescio di colture intercalari o quelle che prevedono lavorazioni minime del terreno, che esercitano forti impatti sulle caratteristiche e sulle fertilità del suolo può essere fortemente condizionata dalla scelta di cultivar dotate di specifiche architetture radicali o di una spiccata plasticità di modulazione di questo tratto morfo-funzionale. Le azioni dimostrative svolte hanno avuto lo scopo di illustrare sperimentalmente questo aspetto.
Figura 1 — Ideotipi di apparato radicale
Le attività a dimostrazione della diversa adattabilità delle cultivar di riso Sole e CL15 all’agrotecnica del sovescio con Vicia villosa in termini di plasticità dell’apparato radicale sono state realizzate presso il campo sperimentale di Nicorvo (PV). Allo scopo si è proceduto alla semina delle due cultivar di riso: a) in due parcelle (3 m x 24 m) distinte, 15 giorni dopo che la veccia su di esse cresciuta era stata interrata; b) in due parcelle delle stesse dimensioni che non erano state occupate dalla coltura intercalare. Trascorsi 30 gg dalla semina (piante allo stadio di IV foglia) si è proceduto a un primo campionamento di radici e subito dopo alla somministrazione di urea (60 kg ha-1 di azoto) e alla sommersione delle parcelle. Trascorsi ulteriori 45 gg è stata somministrata una seconda dose di urea (60 kg ha-1 di azoto). Un secondo campionamento di radici è stato effettuato quando entrambe le varietà erano in fase di fioritura (107 gg dopo la semina).
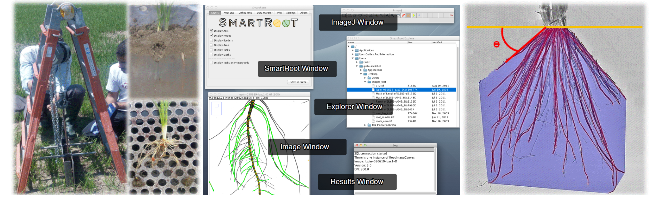
Figura 2 — Campionamenti degli apparati radicali del riso ed analisi della loro architettura
I campionamenti delle radici sono stati realizzati interrando, perpendicolarmente alle fila di semina, due lamine parallele di acciaio dotate di bordi taglienti (Fig. 2a) ed estraendo da esse, una volta rimosse, il blocco di terreno che includeva l’apparato radicale (Fig. 2b). L’eccesso di suolo adeso alle radici era rimosso con prolungati lavaggi (Fig. 2c). Gli apparati radicali erano quindi fotografati e le immagini sono state analizzate con il software SmartRoots® (Fig. 2d). Questo programma permette di definire l’angolo (ϴ) di sviluppo delle radici superficiali rispetto al piano di campagna (Fig. 2e), il numero e la lunghezza media delle radici, nonché la proiezione sul piano del volume occupato dall’intero sistema radicale, cioè l’area dell’inviluppo convesso (AIC; Fig. 2e) definito dall’apparato radicale che stima, con buona approssimazione, il volume di suolo che esso esplora.
Le attività svolte hanno permesso di constatare come nelle prime fasi di crescita delle piante, senza alcun apporto di azoto (N0), né di origine minerale né derivante dalla mineralizzazione della veccia, la cultivar CL15, pur mostrando un numero leggermente inferiore di radici, è caratterizzata da un angolo di crescita radicale e una lunghezza media delle radici superiori a quelli di Sole; ciò si traduce in una esplorazione più profonda e di volumi maggiori di suolo (Fig. 3). La disponibilità di azoto derivante dalla mineralizzazione della veccia interrata ha un impatto evidente sulla architettura radicale di Sole; si riscontra infatti una riduzione (-26%) dell’angolo di crescita, ma un aumento consistente della lunghezza delle radici e del volume del suolo esplorato. Nel caso di CL15, la cui architettura radicale è di per sé più adatta a condizioni di limitata disponibilità di azoto, l’interramento della veccia provoca, coerentemente con una aumentata disponibilità del nutriente, solo una pronunciata riduzione dell’angolo (-38%).
Il secondo campionamento è stato realizzato in fase di fioritura, ovvero quando la coltura aveva ricevuto l’intero apporto azotato (N120) da circa 30 giorni. Sia per CL15 che per Sole, il sovescio determina un aumento della lunghezza delle radici (+17 e +13 % rispettivamente) ed un aumento dell’area esplorata (+ 45 e + 13 %), mentre si è evidenziata una riduzione del numero delle radici per entrambe le varietà. L’analisi dimostra anche che le radici cresciute nelle parcelle interessate al sovescio si sviluppano più in profondità, + 10 e +12 % dell’angolo di crescita, per entrambe le varietà. Ciò è coerente con la rilevata diminuzione di presenza di azoto nel suolo rispetto ai momenti successivi al trattamento fertilizzante e ad una sua probabile lisciviazione verso gli orizzonti più profondi.
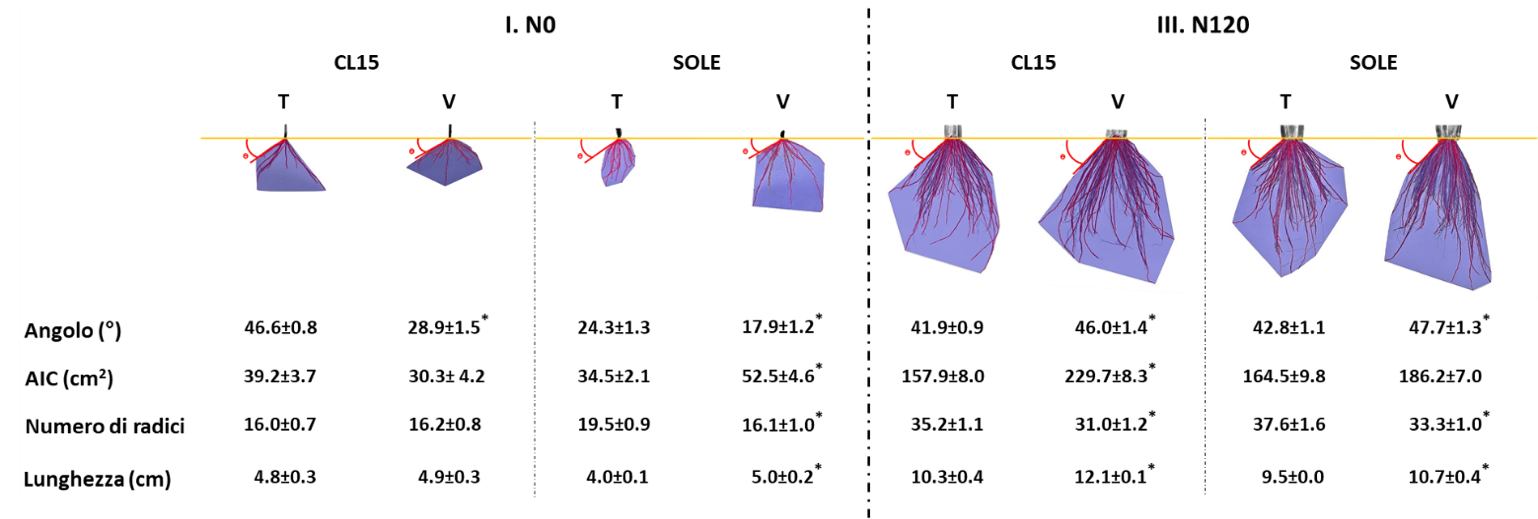
Figura 3 — Cambiamenti dell’architettura radicale delle varietà di riso CL15 e Sole determinati dal sovescio della coltura invernale di V. villosa. I dati riportati rappresentano i valori medi ottenuti nelle parcelle ± ES analizzando venti piante. Gli asterischi indicano l’esistenza di differenze significative (p<0,05) nel confronto tra parcelle interessate (V) o no (T) dal sovescio relativamente ad ognuna delle due cultivar.
Le osservazioni effettuate confermano che:
La seconda parte dell’attività dimostrativa realizzata presso il campo sperimentale di Nicorvo ha avuto lo scopo di illustrare gli effetti del sovescio con V. villosa sull’architettura radicale della cultivar Sole cresciuta a diversi livelli di concimazione azotata (0, 80, 120 kg ha-1 di azoto ureico). In questa attività i campionamenti degli apparati radicali sono stati effettuati 40 giorni dopo l’interramento della leguminosa e in fase di fioritura.
I campionamenti in fioritura hanno mostrato che lo sviluppo delle radici delle piante che sono cresciute solo con l’azoto fornito dalla mineralizzazione della V. villosa interrata è paragonabile a quello osservabile nelle piante fertilizzate con 120 kg ha-1 di azoto; inoltre si osserva come con l’aumento progressivo della dose di fertilizzante, il sovescio determina un’esplorazione del suolo più profonda (Fig. 4). Il sovescio è in grado di influenzare, relativamente alle dosi 0 e 80 kg ha-1 di azoto, anche il numero di radici, riducendolo. L’angolo di inserzione radicale aumenta in presenza di sovescio nel testimone (dose 0 di azoto) ed in presenza di concimazione azotata si rileva anche in questo caso una tendenza ad aumentare l’angolazione.
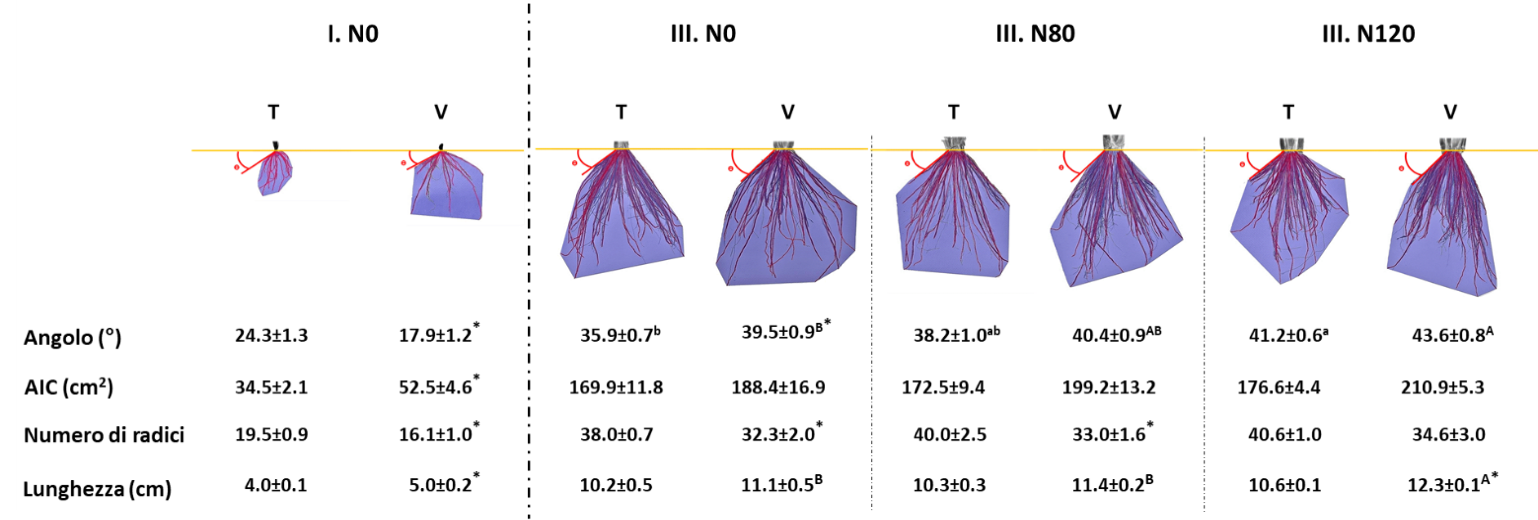
Figura 4 — Cambiamenti dell’architettura radicale delle varietà di riso Sole determinati dal sovescio di V. villosa e da dosi crescenti di urea. I dati riportati rappresentano la media ± SD del valore di ogni parametro misurato in ognuna delle quattro parcelle considerando dieci piante per parcella. Lettere diverse ad apice dei valori indicano, nell’ambito dello stesso trattamento (testimone, T o sovescio, V), l’esistenza di differenze significative (p<0,05; n=4) in funzione di apporti crescenti di fertilizzante ureico. Gli asterischi indicano invece l’esistenza di differenze significative (p<0,05; n=4) tra parcelle che hanno ricevuto la stessa dose di fertilizzante, ma che sono state interessate o meno dal sovescio.
Contemporaneamente al campionamento delle radici si è proceduto alla valutazione dello stato nutrizionale azotato delle piante di Sole cresciute nelle parcelle interessate al sovescio e a diverse dosi di fertilizzante ureico utilizzando il sensore ottico Dualex®. Questo strumento (Fig. 5a) rileva in modo rapido e non distruttivo i livelli di clorofilla e flavonoidi presenti in una foglia traendo dal loro rapporto un indice, definito Nitrogen Balance Index (NBI), che esprime lo stato azotato della pianta.
Nelle foglie delle piante cresciute per 30 giorni senza alcun apporto di azoto (testimone) o solo con quello apportato dalla mineralizzazione dalla veccia interrata (sovescio), i valori di NBI sono del tutto simili (Fig. 5b). Ciò indica come in questa fase di sviluppo la necessità di azoto delle piante sia soddisfatta dalla dotazione naturale del suolo. In fase di fioritura invece per raggiungere il valore massimo di NBI è necessaria la dotazione di azoto fornita da 160 kg ha-1 di urea, mentre nel caso delle parcelle interessate al sovescio ciò è raggiunta a dosi minori del fertilizzante azotato (Fig. 5b).
Le osservazioni effettuate confermano:
In mais si stanno sviluppando modelli molecolari che possono fornire indicazioni rapide sullo stato azotato della coltura che possano affiancarsi ai metodi chimici tradizionali. Tra questi sembra promettente un sistema che si basa sull’analisi combinata degli andamenti d’espressione di alcuni geni specificamente sensibili allo stato del metabolismo azotato della pianta. Al fine di confermare anche in riso la validità di questo approccio si è proceduto alla analisi di espressione dei geni (Os05g49240, Os09g37710, Os08g38700 e Os03g07570) ortologhi a quelli di mais identificati come utili. I valori di espressione relativa dei quattro geni sono stati correlati ai valori di NBI mostrati dalle piante della varietà Sole cresciute sino alla fase di inizio accestimento nelle parcelle a diversa disponibilità di azoto derivante dagli apporti di fertilizzante minerale e/o dalla mineralizzazione della veccia interrata.
Figura 5 — Stato nutrizionale azotato (valori di NBI) delle piante della cultivar Sole cresciuta in parcelle interessate o meno al sovescio con V. villosa e fertilizzate con dosi crescenti di urea. Le barre dell’istogramma rappresentano la media ± ES del valore di NBI misurato in ognuna delle quattro parcelle considerando dieci piante per parcella. Lettere diverse indicano nell’ambito dello stesso trattamento (testimone o sovescio) l’esistenza di differenze significative (p<0,05; n=4) in funzione di apporti crescenti di fertilizzante ureico. Gli asterischi indicano invece l’esistenza di differenze significative (p<0,05; n=4) tra parcelle che hanno ricevuto la stessa dose di fertilizzante, ma che sono state interessate o no dal sovescio.
L’andamento dei profili d’espressione di questi geni è, in modo del tutto simile a quanto dimostrato in mais, coerente con lo stato azotato delle foglie: la loro espressione diminuisce al migliorare dello stato nutrizionale azotato delle piante e tra le due variabili esiste una relazione quantitativa (Fig. 6), dimostrando come i geni analizzati risultano quindi idonei per valutare, con buona precisione, anche lo stato nutrizionale azotato del riso in tempo reale.
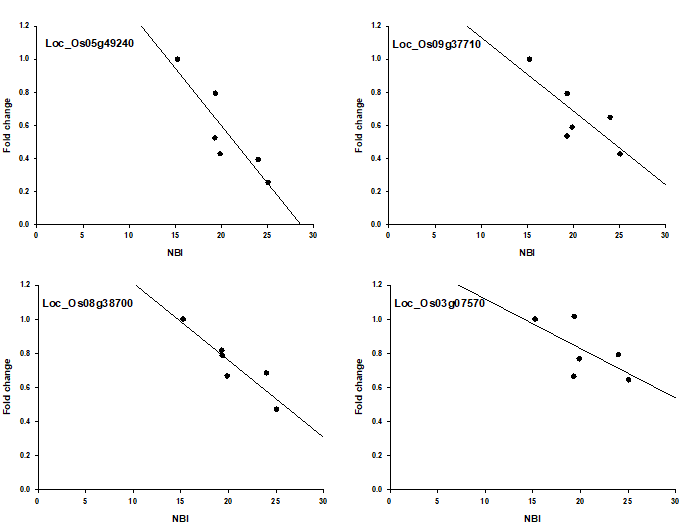
Figura 6 — Relazione tra i valori di NBI e i livelli di espressione di quattro geni specifici nelle foglie della cv Sole cresciuta in diversa disponibilità di aoto nel suolo.
Le osservazioni effettuate confermano:
L’influenza della pratica del sovescio e delle lavorazioni conservative sulle popolazioni microbiche rizosferiche delle varietà Sole e CL15 è stata valutata mediante conte di batteri rizosferici aerobi ed anaerobi totali ed azoto-fissatori.
Al momento della semina e dopo 60 giorni dalla semina, sono state campionate rizosfere di piante di riso cresciute su terreni sovesciati. Nelle due varietà testate e in tutte le condizioni agronomiche, il numero di microrganismi totali e azoto-fissatori aumentava nel corso della stagione vegetativa.
Come mostrato in Fig. 7 il titolo dei batteri aerobi totali della varietà Sole nella coltura sovesciata o non sovesciata non variava. Quello della varietà CL15 era inferiore nella coltura sovesciata rispetto alla coltura non sovesciata. In entrambe le varietà il sovescio promuoveva un incremento di batteri anaerobi. Il numero dei batteri azoto-fissatori, sia aerobi che anaerobi, era inferiore in condizioni di sovescio per entrambe le varietà.
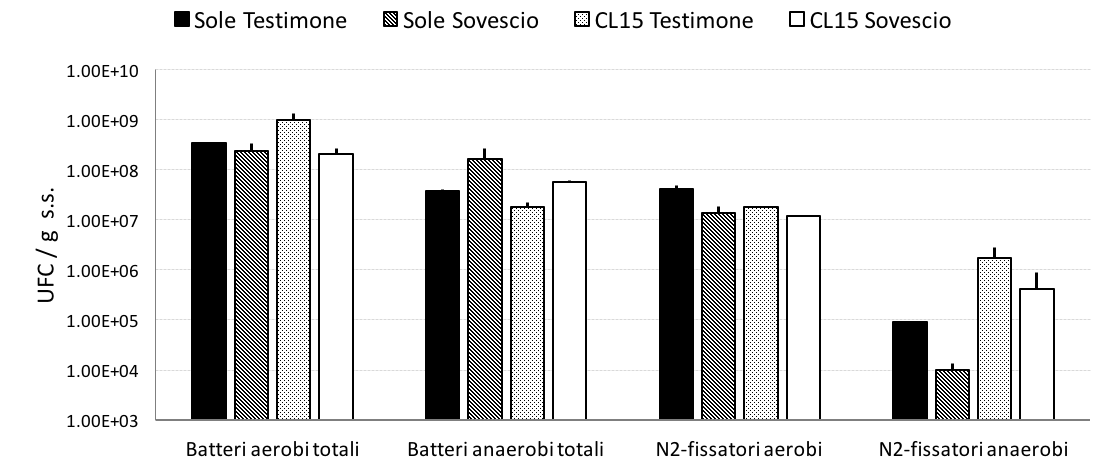
Figura 7 — Incremento del titolo di batteri rizosferici (calcolato come differenza tra titolo batterico dopo 60 giorni dalla semina e al momento della semina) delle varietà Sole e CL15, nelle condizioni di suolo non sovesciato (testimone) e sovesciato.
Le osservazioni effettuate permettendo di concludere che:









